


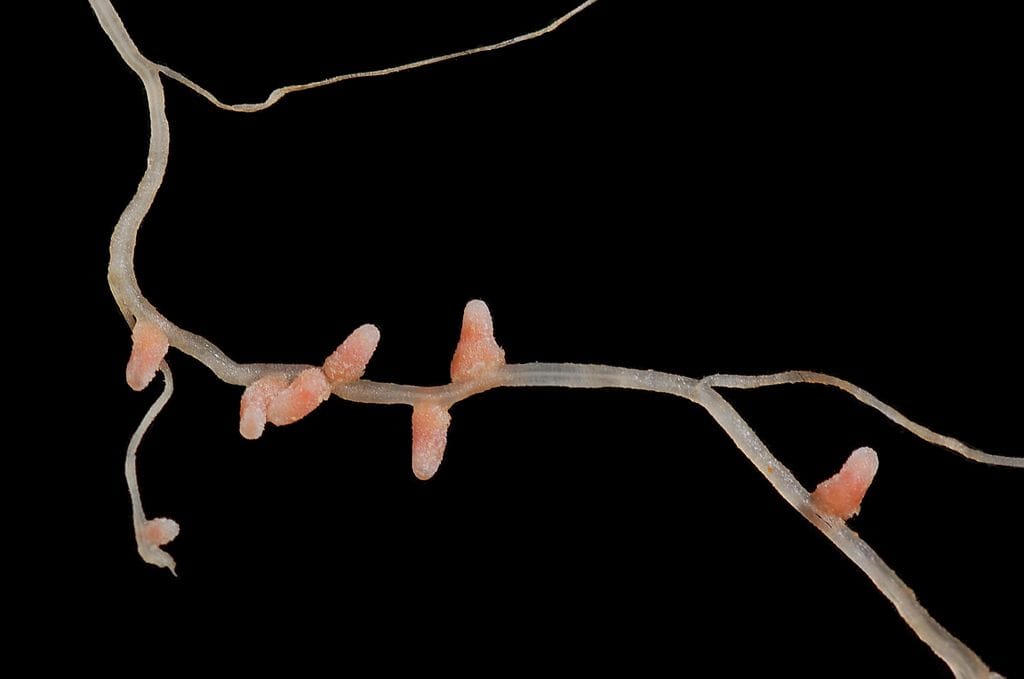
The legume family of plants can form a symbiosis with nitrogen-fixing bacteria known as rhizobia. In return for a supply of nitrogen compounds the plant provides sugars and houses the rhizobia in special organs called nodules (see figure). The development of the nodules and the infection of rhizobia into them are tightly regulated by the plant. Many genes are required for the establishment of a successful symbiosis so how did it evolve?
The simple answer is that many of the genes were co-opted from their existing functions in symbioses with mycorrhizal fungi. The mycorrhizal symbiosis is ancient, appearing around 450 MYA when plants were first colonising the land. Today, around 90% of land plants including the legumes can form mycorrhizal associations, with the plant receiving nutrients including phosphates and nitrogen-compounds (and other benefits) in exchange for providing sugars (1). The rhizobial symbiosis is much more recent (about 60 MYA) and is almost exclusive to the legumes.
Both symbioses share an early signalling pathway, called the common symbiosis (or Sym) pathway. Central to this pathway are calcium oscillations or “spiking” generated and decoded by a set of shared genes (highlighted in yellow in the figure below).
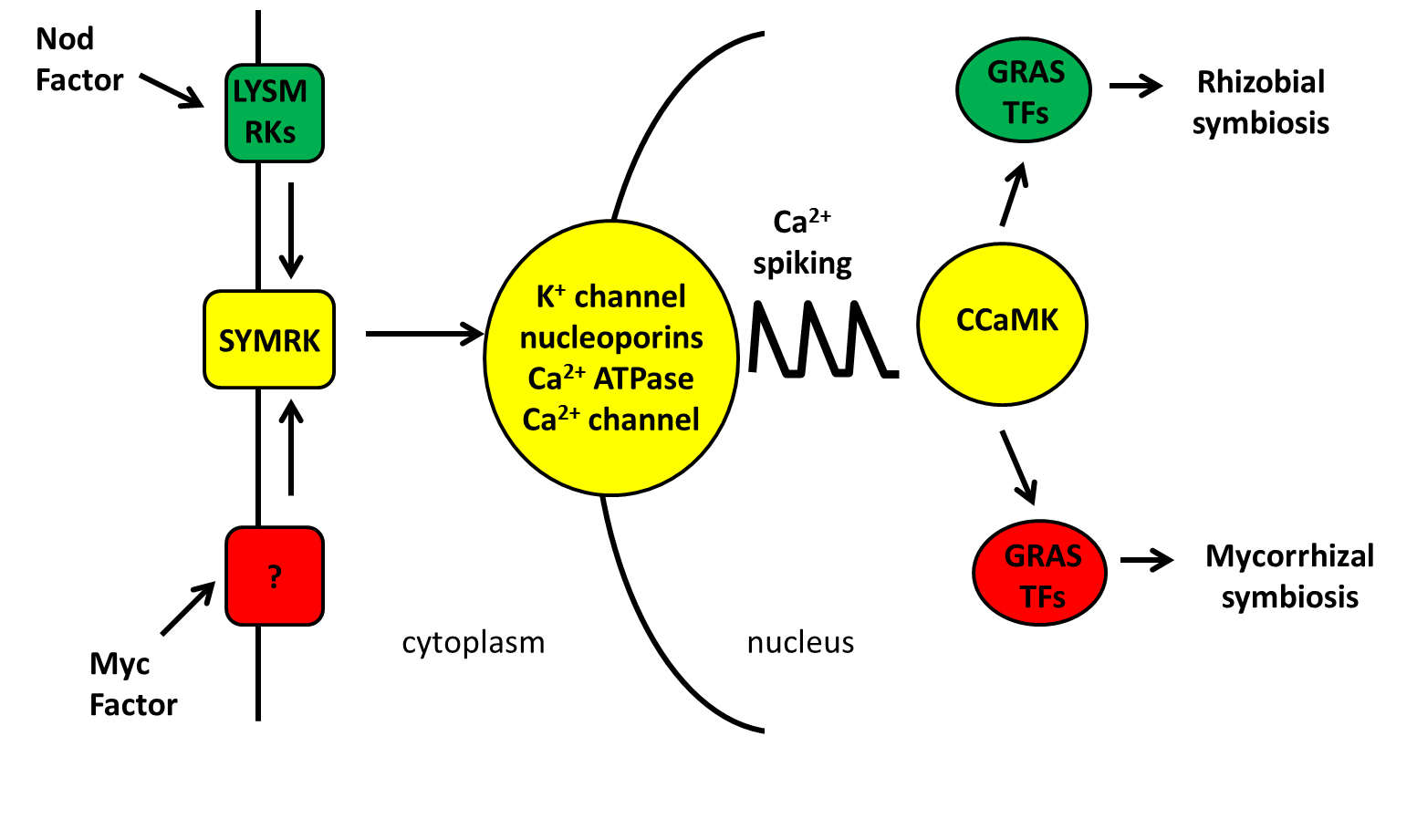
Downstream of calcium spiking the rhizobial and mycorrhizal signals split leading to the different changes in gene expression required for each symbiosis. Both symbioses require members of the GRAS transcription factor family and it appears that while some members are utilised in both symbioses, other members are specific for one or the other.
The similarities are not restricted to within the plant. Both symbioses start with an exchange of signals between the plant and the microbe. The signals produced by the mycorrhizal fungi (Myc factors) and the rhizobia (Nod factors) are actually very similar, being chitin chains with various lipid modifications known as known as Lipochitooligosaccharides (LCOs). Chitin is the main constituent of fungal cell walls but bacteria don’t usually make chitin so how rhizobia developed the ability is unclear. It is possible that rhizobia acquired the genes required for LCO synthesis from fungi by horizontal gene transfer. Alternatively, Nod factor production may have evolved separately by altering existing bacterial metabolic pathways (peptidoglycan, the major component of bacterial cell walls is structurally similar to chitin).
The receptors that perceive Nod factors belong to the LysM receptor-like kinase family. Other members of this family include a chitin receptor (CERK1) involved in defence against fungal pathogens. The Myc factor receptors are still unknown but it seems likely that they belong to this family too.
So, rhizobia became able to mimic mycorrhizal fungi by producing LCOs and the plant modified existing signalling pathways. Plant-guided rhizobial infection into the root is also thought to have been adapted from mycorrhizal fungi infection. This opens the possibility of genetically modifying non-legume crop plants to form the nitrogen fixing symbiosis with rhizobia. The major cereals (rice, maize and wheat) can form mycorrhizal symbioses so in theory only the genes from legumes that modify the existing signalling pathways to mediate rhizobial infection plus those required for nodule development would need to be introduced. The ENSA project, a collaboration between several labs in Europe and the US are attempting to engineer cereals to be able to perceive rhizobial Nod factors and respond with some basic gene expression changes. Nitrogen-fixing cereals are still a way off but it’s a start.
References
Kistner C. & Parniske M. (2002). Evolution of signal transduction in intracellular symbiosis,
Trends in Plant Science, 7(11) 511-518. DOI: 10.1016/S1360-1385(02)02356-7
Images
Figure 1. Medicago truncatula root with nodules infected with Nitrogen-fixing rhizobia by Ninjatacoshell/Wikpedia. [cc]by-sa[/cc]
Figure 2. Common sym pathway. Image by author.




{kind=link}