


Arguably, one of the best known ‘structure-function’ relationships in plant biology is the role played by cellulose microfibrils within the walls of the guard cells in stomatal opening. Stomata* are the controllable orifices found primarily within the epidermis of the above-ground organs of higher plants. When open they permit ready exchange of gases (e.g. CO2, O2) between the interior of the plant and the environment thereby facilitating photosynthesis, and aerobic respiration. However, when open, H2O – in the gaseous form of water vapour – can also leave the plant in the process known as transpiration.
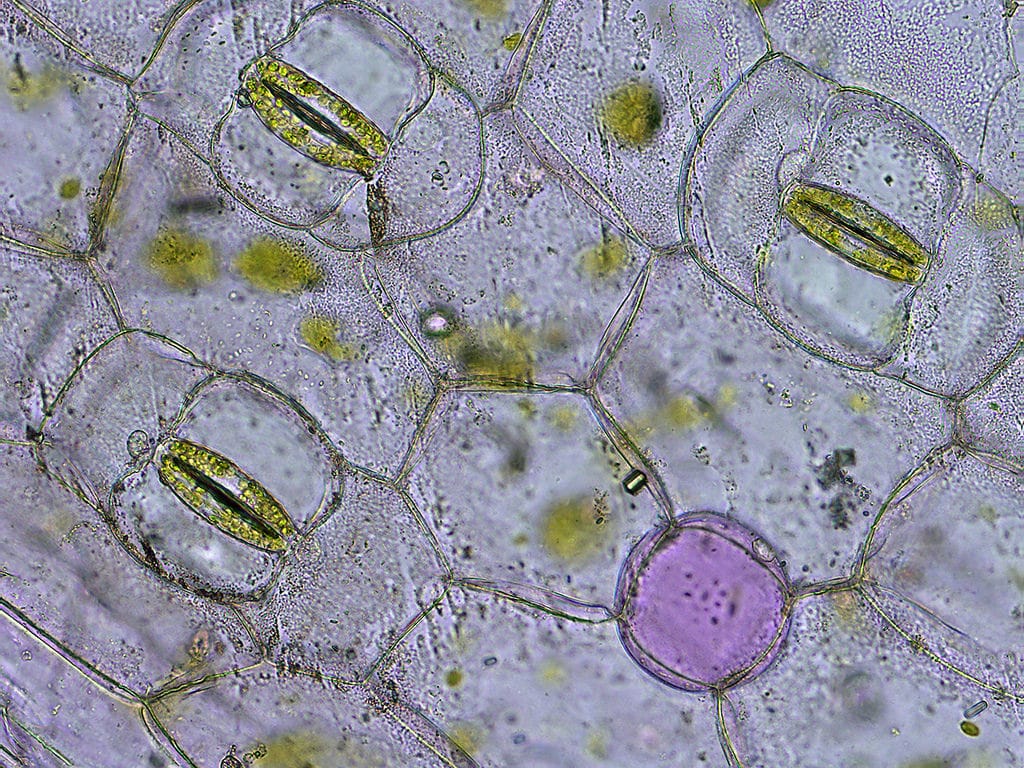
Where water in the soil is in sufficient supply, transpirational water loss is a small price to pay for ready uptake of photosynthetically-essential CO2 via the open stomata**. Whilst stomatal opening is ultimately down to the uptake of water into the vacuoles of the pair of guard cells (the cellular components that border the stoma) and which therefore become turgid, it is the arrangement of the microfibrils of cellulose within their cell walls that ensures that the opening is properly formed. Cellulose microfibrils resist stretching and compression in the direction parallel to their orientation. Since those microfibrils are arranged as ‘hoops’ around the circumference of the guard cells, this constraint permits only increase in length when the cells are turgid. However, since the guard cells are attached to each other at their tips, the increase in length causes the cells to buckle, and separate. This ‘bowing-apart’ of the guard cells generates the stomatal pore and is a natural consequence of their increasing more in length than width as turgor increases.
Like many people I suspect I presumed those cellulose microfibrils were set in place once and stayed there throughout the life of the guard cell, and was perfectly happy to leave matters there. Fortunately, and unlike most other people, Penn State University (USA) ‘stomatologists’ Yue Riu & Charles Anderson didn’t leave matters there. Examining the roles of cellulose and xyloglucans (XG) in Arabidopsis guard cell walls they reveal a dynamic system in which those two major cell wall components interact during stomatal opening/closure (Plant Physiology). Importantly, they demonstrate that cellulose microfibrils undergo dynamic reorganization during stomatal movements.***
Furthermore, using plants deficient in cellulose – the cesa3je5 mutant (e.g. Andrew Carroll et al., Plant Physiology 160: 726-737, 2012) – they demonstrated that greater stomatal opening than in wild type plants could be achieved (!), apparently because changes in guard cell length occur more rapidly in such individuals. So, rather than facilitate maximal stomatal aperture, cellulose actually appears to constrain this. Who’d’ve thought?
The interplay between XG and cellulose also revealed here is seemingly just another example of a more widespread interaction between these two cell wall components that impacts upon aspects of plant cell growth and morphogenesis more generally (Chaowen Xiao et al., Plant Physiology 170: 234-249, 2016). Which only serves to remind us that as static as cell walls may appear there is an awful lot going on inside them!
* Final Year Philosophical Botany exam question: “Stomata the most important orifices on the planet: Discuss”…
** Where water is not so abundant, and can’t replace that lost by transpiration, conservation of this essential life-giving fluid that is already within the plant, is achieved by stomatal closure; prevention of further loss of water therefore takes priority over photosynthetic-promoting uptake of CO2.
*** How many opening/closing cycles can any stoma undergo before it is ‘worn out’? If it has a finite ‘lifespan’, and it ‘expires’ before the death of the organ it is embedded within, does it remain open or closed, or somewhere in between? Does that then pose a risk of uncontrollable water loss for the plant..?


