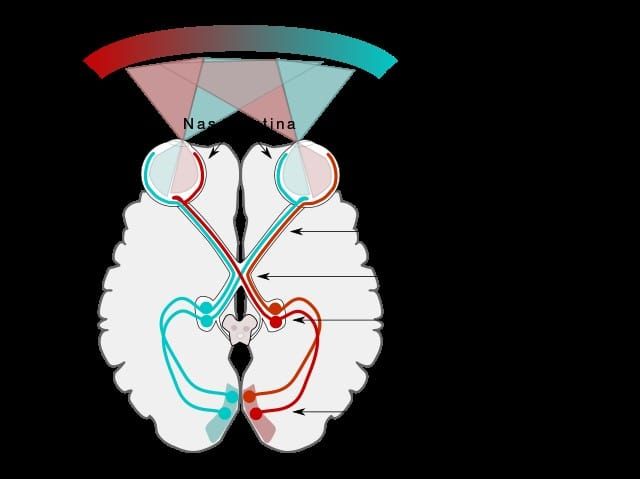
With all the hi-tech, whizzy methodology at our disposal nowadays you can be forgiven for thinking that we know all we need to know about plants. But, and I always find this realisation rather humbling, that is not the case; there is still a great deal we don’t yet know. And one of the most enduring mysteries has been the role(s) played by calcium oxalate deposits in plant tissues.
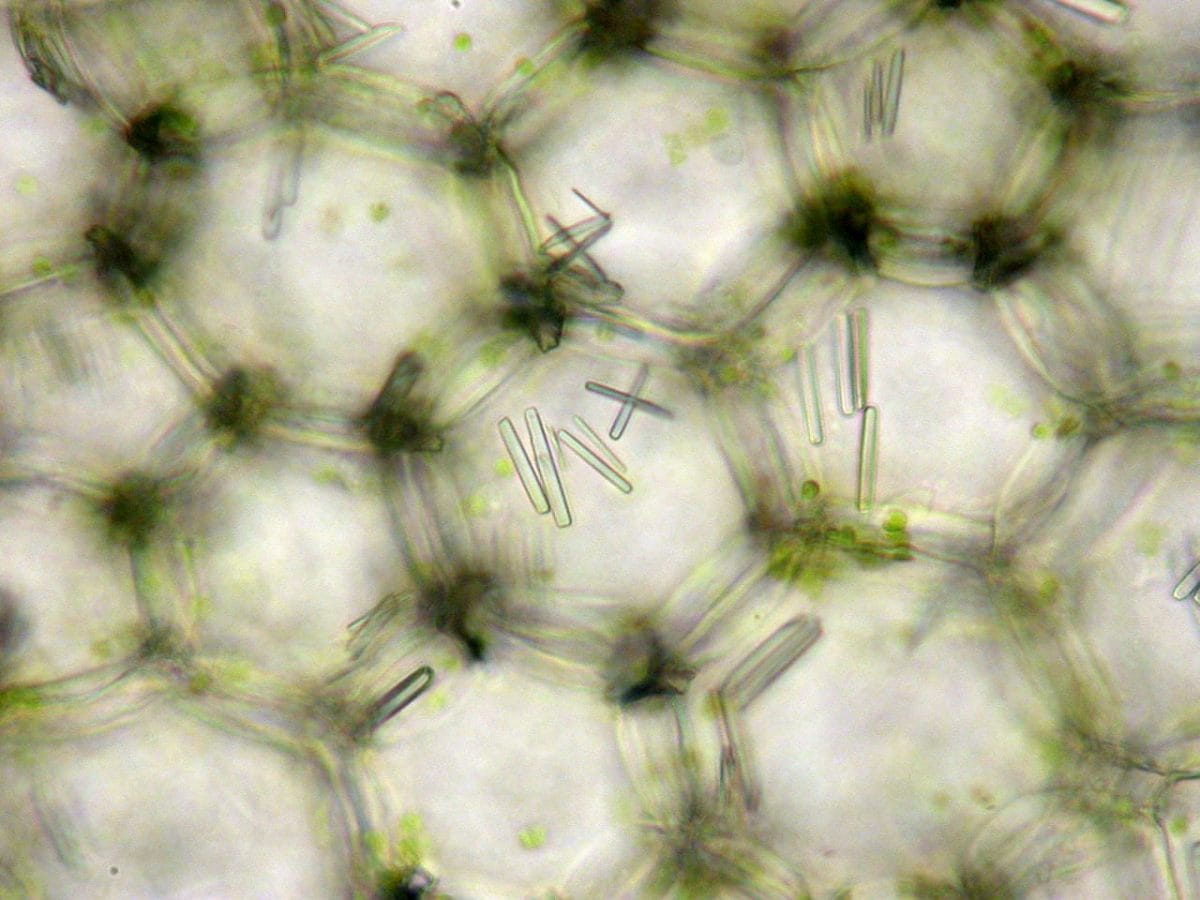
Due to the fact that these mineral inclusions contain calcium, they have been posited as a store of that essential plant macronutrient (which has assumed additional importance as a so-called second messenger – whose concentrations increase dramatically in response to a wide range of abiotic and biotic phenomena and which thereby provides a link between perception of environmental change and the molecular genetic events that permit the plant to respond appropriately). And, because they also contain oxalic acid, which is poisonous in appropriate concentrations, they have assumed herbivory-defence (e.g. this) or human-irritant roles, in addition to their calcium storing function. Furthermore, the crystals’ size and frequently sharp-pointed nature have also suggested a more physical deterrent to would-be consumers of the plant tissue that houses these crystalline creations.
But, what person(s) would have looked beyond this ‘obvious’ bipartite chemical nature and pondered the fact that CO2 is also effectively a constituent of oxalic acid? Georgia Tooulakou et al., that’s who.
In their fascinating study they propose the exciting notion that these crystals provide an internal source of CO2 for plants. Examining mesophyll cells of pigweed (Amaranthus hybridus), they noticed daily fluctuations in crystal volume – which decreased during daylight but recovered at night-time. Decrease in volume was due to crystal decomposition, associated with increased activity of oxalate oxidase, an enzyme that converts oxalate into CO2 and H2O. Intriguingly, isotope analysis established that the carbon in the oxalate was derived from a non-atmospheric CO2 source. And, despite stomata being closed after midday in A. hybridus (presumably a water-saving strategy that limits transpirational water loss in response to droughted conditions, and which would otherwise effectively prohibit any sustained photosynthesis in this C3 plant by preventing entry of atmospheric CO2), there is evidence that photosynthesis continues to take place.
Putting all this together, the team propose that leaf-localised calcium oxalate crystals act as a biochemical reservoir that stores non-atmospheric carbon (derived e.g. from respiration), mainly at night, and which is enzymatically-released inside the plant during the day for photosynthetic assimilation, particularly under drought conditions. [Ed. – and the water produced by oxalate-oxidation is presumably also useful under these conditions.]
Thus, A. hybridus – nominally, a C4 photosynthesis plant – appears to have developed an elegant alternative to CAM (Crassulacean Acid Metabolism – the variant of photosynthesis whereby stomata are open at night-time when atmospheric CO2 is fixed into 4-carbon organic acids and subsequently released internally during daylight – when stomata are closed – which permits its reincorporation into sugars via C3 photosynthesis). Plants, ever-resourceful survivors whatever the environment throws at them. Isn’t nature wonderful? [Clue, the answer is YES!]
*This article refers to the mineral deposits known as druse crystals or raphides. If you are interested in the religious sect known as the Druse, you need to look elsewhere.
[Ed. – this hypothesis, so-styled ‘alarm photosynthesis’, is further explored in ‘Reevaluation of the plant “gemstones”: Calcium oxalate crystals sustain photosynthesis under drought conditions’ by Georgia Tooulakou et al.. And for some semblance of balance, to find out more about the role of silica-based mineral inclusions in plants, phytoliths, we recommend Caroline Strömberg et al..]