


The top of a mountain is a bit like an island: high elevation mountain habitats are surrounded by a sea of radically different lowland environments, which isolate the mountain habitats from one another and can act as a barrier for the dispersal of species. However, despite these barriers, some mountain habitat “islands” can share quite similar flora. For example, amongst the various conifers native to Africa, the Podocarpus latifolius – Podocarpus milanjianus complex is found on mountaintops throughout the Eastern, Central and Southern African highlands. With a “sea” of lowland environments between these various mountain-top “islands”, the question arises: how did the P. latifolius – P. milanjianus complex manage to become so incredibly widespread?

A recent study by Migliore and colleagues set out to find an answer. The team collected leaf samples from each part of the conifer’s range, from Cameroon to Angola in western Central Africa and from Kenya to the Cape region in eastern and southern Africa. For each sample, they sequenced the whole plastid genome, mitochondrial DNA regions, and nuclear ribosomal DNA, to figure out the phylogenetic and demographic history of the P. latifolius – P. milanjianus complex.
Their first major finding was that P. latifolius and P. milanjianus are actually the same species – the two names have been thought to represent just one species for quite some time, but Migliore and colleagues finally confirmed it. They wrote: “as South-African samples (P. latifolius) did not form a clade separated from the other ones (P. milanjianus), we confirm that the two taxa can be considered as synonyms”. This initial discovery makes the widespread distribution of P. latifolius/milanjianus even more interesting. How did this single species end up on so many different isolated mountain habitats?
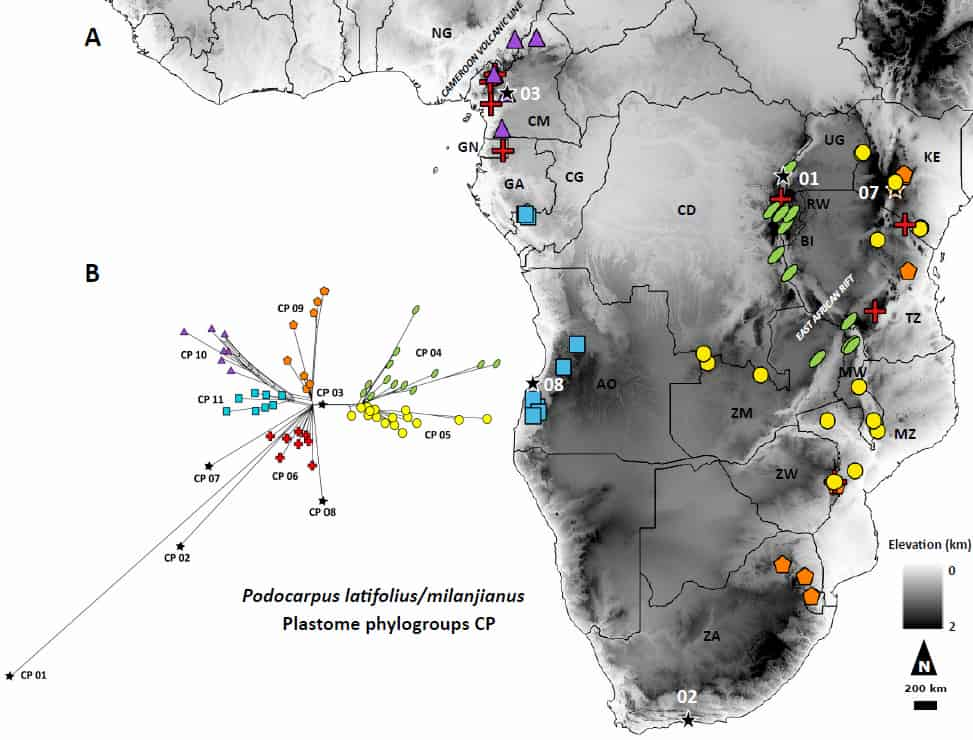
Migliore and colleagues argue that the various mountain habitats where P. latifolius/milanjianus currently occurs were better connected in the past. They found that the P. latifolius/milanjianus lineages “diverged during the Pleistocene, mostly between c. 200 and 300 kyrs BP, after which they underwent a wide range expansion leading to their current distributions”. Based on phylogenomic and palaeoecological data, they argue that the species originated in East Africa, and then spread through two probable migration corridors: a northern route towards the Cameroon volcanic line, and a southern one towards Angola. Unfortunately, the distribution of P. latifolius/milanjianus is extremely fragmented and the species is unfortunately now rare in large parts of its range. Migliore and colleagues suggest that after the species’ range expansion during the Pleistocene, it must have experienced a demographic decline in large parts of its range. While they couldn’t detect this in their DNA sequences, the authors suggest that the decline may have occurred too recently to have left a genetic signature.
The authors conclude that “a long-standing question in African biogeography is whether colonisation of the mountains and subsequent intermountain gene flow mainly depend on long-distance dispersal across unsuitable lowland habitat or on intermittent suitable habitat bridges”. Their research on Podocarpus latifolius/milanjianus suggests the latter: that the montane forest habitats extended to lower elevations in the past, which allowed for widespread species migration over habitat bridges between the mountain-top islands.





{kind=link}